Qu'est ce qu'un Stimulus
La réaction musculaire ou nerveuse de l’homme dépend d’une excitation entraînée par un stimulus. Ainsi, un stimulus peut être défini comme étant le facteur déclenchant une réponse physiologique. Il peut prendre plusieurs formes:
- un stimulus visuel lorsqu’il s’agit d’une réaction à une image
- un stimulus auditif lorsqu’il s’agit d’une réaction à un son
- un stimulus tactile lorsqu’il s’agit d’une réaction à une pression sur la peau
- un stimulus thermique lorsqu’il s’agit d’une réaction à un changement de température
- un stimulus olfactif lorsqu’il s’agit d’une réaction à une odeur
Comment notre Corps le Détecte?
L’œil réagira à une onde électromagnétique du spectre visible, l’oreille à une vibration mécanique dans l’air ou à une pression sur la peau, et le nez à des molécules odorantes dans l’air. Le mécanisme de réaction face à un stimulus n’est pas le même dans tous les cas, différents récepteurs sont sensibles à des stimuli de natures différentes:
- les photorécepteurs répondent aux longueurs d’ondes dans le spectre visible
- les mécanorécepteurs répondent à de l’énergie mécanique, tel des vibrations dans l’oreille ou une pression exercée sur la peau
- les chimiorécepteurs répondent à des éléments chimiques spécifiques (les cellules réceptrices de la cavité nasale ne vont pas réagir aux mêmes molécules que les papilles)
- les thermorécepteurs répondent au chaud et au froid
- les nocicepteurs répondent aux agressions pouvant endommager les tissus. Une stimulation excessive de ces récepteurs est perçue comme de la douleur.
Expérience: Rapport entre stimulus auditif inattendu et rythme cardiqaue
Objectif
Observer le temps de réaction grâce à la réaction d'un muscle (le cœur) face à un stimulus auditif inattendu (claquement des mains).
Matériel
- ordinateur
- logiciel Oscillo 5
- l'interface d'aquisition
- cardio-fréquencemètre
- microphone
Protocole
On branche l'interface d'aquisition à l’ordinateur, puis le cardio-fréquencemètre et le microphone à l'interface d'aquisition. Ensuite on ouvre le logiciel “Oscillo 5”. On choisit les paramètres adaptés pour avoir des bons graphiques. On demande aux personnes testées de poser leur pouce sur le cardio-fréquencemètre afin d’obtenir une courbe de la fréquence cardiaque au repos sur l’écran. En gardant le pouce sur l’appareil, une autre personne claque des mains à un moment inattendu. Le microphone enregistre le son du claquement des mains afin de voir la réaction par rapport au stimulus sur le graphe. On observe la fluctuation de la courbe sinusoïdale pour en tirer des conclusion.
Résultats
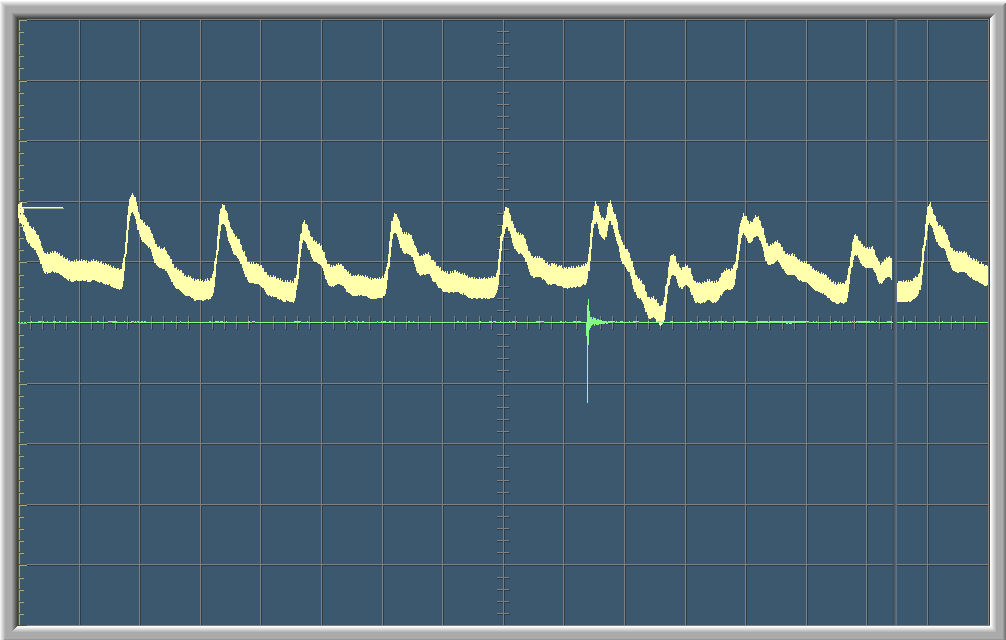
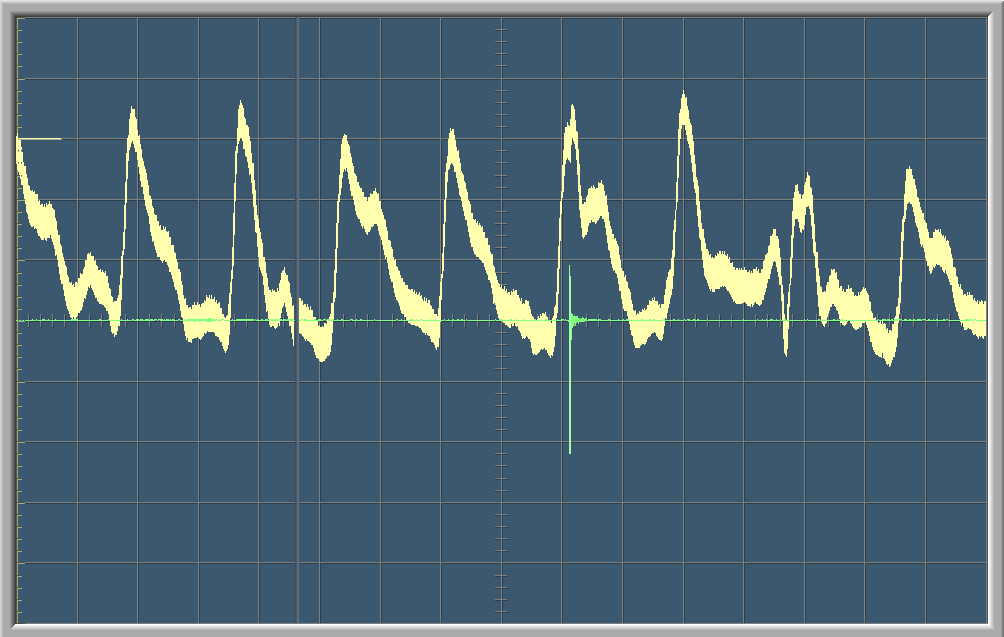
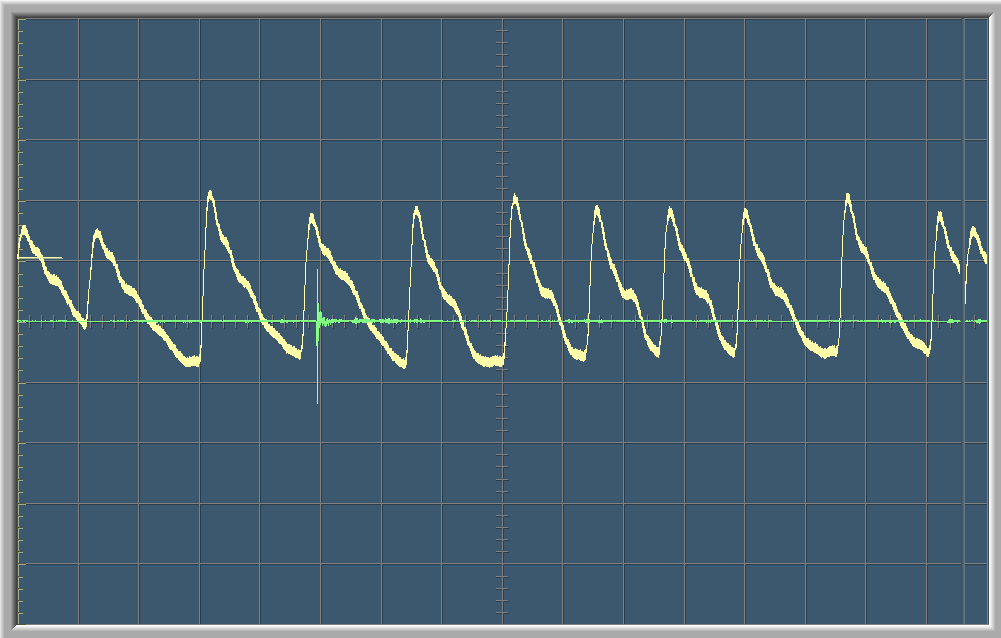
Commentaires et Calculs
La courbe verte sur le graphique montre les ondes sonores enregistrées par le microphone. Le pic sur la courbe vert marque le moment qu’on a claqué des mains. La courbe blanche est une visualisation graphique de la fréquence cardiaque. Sur les deux premiers graphes on aperçoit une courte variation dans la fréquence cardiaque de la personne testée au moment du stimulus auditif. Après un court moment, la fréquence cardiaque redevient normal.
Chaque graphe à un balayage de 8 secondes, c’est-à-dire que l’écran se renouvelle toutes les 8 secondes.
Sur le premier graphe, on peut compter une période T=0.8. Grâce à l’équation qui lie la période et la fréquence, nous pouvons en déduire la fréquence:f=1 / T
Sur le deuxième graphe, on peut compter une période T=0.85.
Sur le troisième graph on peut compter une période T=0.95.
Conclusion
Nous avons observé sur deux des trois cas, la fréquence cardiaque augmente grandement après le stimulus. Le coeur se remet alors à son rythme normal après un ou deux battements après que l’individu s’aperçoit que rien de dangereux n’est présent. On en conclut qu’il y a un temps de réaction presque immédiat après le stimulus auditif inattendu, qui se traduit par une augmentation de la fréquence cardiaque. On ne peut pas mesurer exactement le temps que prend le coeur à réagir car l’écran n’est pas assé précis. On peut seulement observer que le coeur prend environ 200ms à réagir au stimulus auditif. Étant donné qu’on observe aucune différence de fréquence cardiaque dans le dernier essai, on peut supposer que le corps n’était pas effrayé dans ces conditions. Par contre, il semble qu’entre les battements, après le claquement, le corps à eut le temps d’annuler la réaction attendu car il a compris que le stimulus n’a rien de dangereux. L’analyse complète du stimulus prendrait donc moins de 500ms alors que la réaction spontané prendrait environ 100ms afin de stimuler une réaction cardiaque. Cependant, il est aussi possible que ce stimulus n’a eut aucun effet sur la personne testée dans le troisième graphe ce qui expliquerait pourquoi celle-ci ne montre aucune réaction au niveau du muscle cardiaque après le stimulus.
Nous avons mis en évidence la réaction du muscle après un stimulus uaditif. Comment se fait cet acheminement, du stimulus au muscle?
Le parcours du stimulus dans notre corps, de l’oeil/oreille au muscle
Pour produire une action face à un stimulus, plusieurs étapes s’enchaînent entre le contact du stimulus et le mouvement du muscle. Le temps de réaction se décline en plusieurs étapes: l’atteinte du capteur par le stimulus, l’envoi de l’information au cerveau, le transfert de l’information du cerveau aux muscles et finalement le déroulement de l’action. Nous allons détailler chaque étape.
L’atteinte du capteur par le stimulus et l’envoi de l’information au cerveau
De l’oeil au cerveau
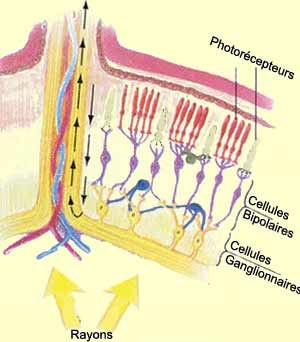
Schéma de l'anatomie de la rétine
Lorsque la lumière pénètre la cornée, elle va traverser l’iris qui, par une dilatation ou un relâchement, contrôle l’intensité de la lumière qui passe par la pupille. Puis, la lumière sera courbé par le cristallin afin d’être envoyé sur la rétine, qui agit comme un film photographique. En effet, la rétine est constituée, en partie, par des photorécepteurs qui captent les signaux lumineux afin de les transformer en signaux électriques: c’est la transduction du signal lumineux. Puis, le signal électrique est envoyé au cerveau via le nerf optique.
Nous avons effectué la dissection d’un oeil de boeuf afin d’illustrer nos propos:
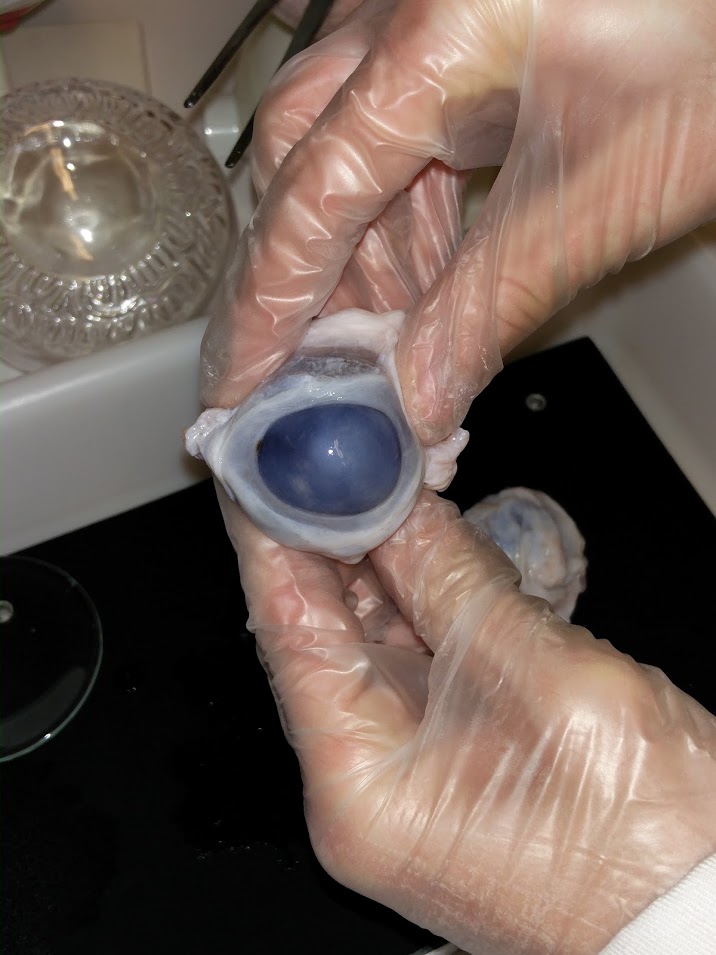
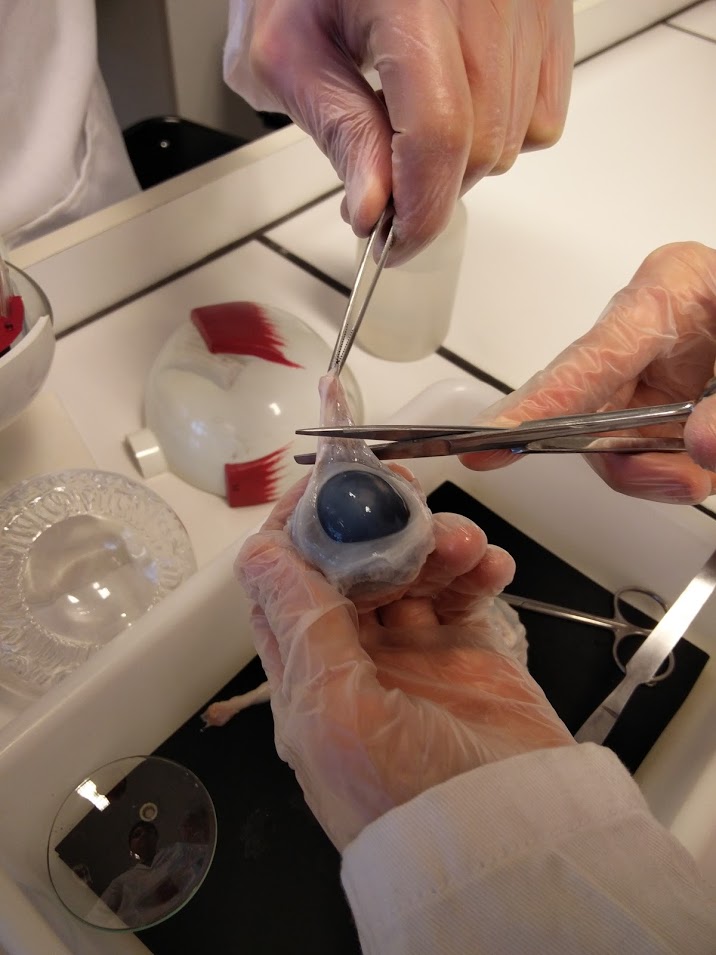




Le sens du traitement dans la rétine est contraire au sens de la lumière. Ainsi les photorécepteurs se trouvent à l'extrémité de la rétine opposé à l'arrivée de la lumière. Le photorécepteur est composé de trois parties: le segment externe, où se trouvent des disque qui vont absorber la lumière, le segment interne qui accueil les éléments nécessaire à la transformation du signal lumineux en signal biochimique, et une terminaison synaptique qui libère le glutamate (lorsque le photorécepteur est hyperpolarisé).
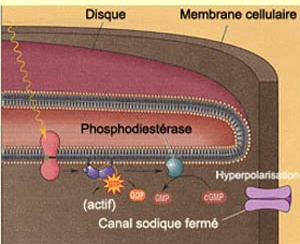
Schéma de la fermeture des canaux à sodium
Le segment externe est composé de disques où siège les éléments nécessaires à la transformation du signal lumineux. En cas d’obscurité, des canaux de sodium situés en extrémités des photorécepteurs sont maintenus ouverts grâce à une production continu de GMPc. Lorsque ces canaux sont ouverts, des ions sodium peuvent entrer au sein de la membrane du photorécepteur. Des canaux de fuites situés au niveau du segment interne du photorécepteus permettent au sodium de sortir de la membrane du photorécepteur. Ce courant continu d’ions sodiums s’appelle le courant d’obscurité, et lorsque les canaux de sodium sont maintenus ouverts, il y a une différence faible entre le potentiel intra et extra-cellulaire.
Le photorécepteur est dépolarisé et produit le neurotransmetteur glutamate. Lorsque la lumière rentre en contact avec un pigment visuel du disque, une cascade de réactions chimiques se déroule: la rétinène, contenu dans une protéine enzymatique (opsine), va changer de conformation. Ce changement mène à l’activation d’une protéine G qui libère sa fraction alpha. Cette fraction alpha active l’enzyme phosphodiétérase, qui élimine les messagers GMPc, et par conséquent, ferme les canaux à sodium sur le segment externe. Puisque les ions ne peuvent qu’échapper du photorécepteurs, il y a une augmentation de potentiel intra et extra-cellulaire.
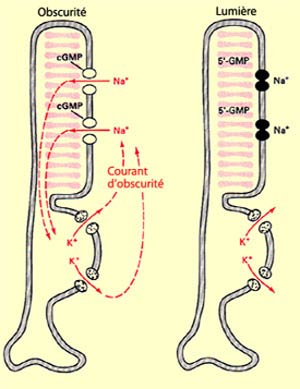
Schéma du courant d’obscurité
Le photorécepteur est hyperpolarisé, et ne relâche plus de glutamate. La particularité des photorécepteurs est que le stimulus l’inhibe, tandis qu’il est excité en absence de lumière.Plusieurs photorécepteurs sont reliés à une unique cellule bipolaire. Cette cellule bipolaire à pour fonctionnement de traiter au préalable l’information reçue par le photorécepteurs (c’est pour cela que l'oeil peut-être considéré comme un prolongement du cerveau). L’ensemble de photorécepteurs auquel une unique cellule bipolaire est lié s’appelle son champ récepteur, qui contient une partie centrale, où les photorécepteurs transmettent directement de l’information aux neurones bipolaires, et une partie périphérique, où des cellules dites horizontales transmettent l’information aux neurones bipolaires. Un photorécepteur du champ récepteur va relâcher plus ou moins de glutamate selon l’endroit sur le champ récepteur où la lumière atterrit. La réponse de la cellule bipolaire dépend également de quel type de cellule bipolaire il s’agit. Il en existe deux types: à centre ON (le relâchement de glutamate au niveau centrale mène à une dépolarisation, soit l’excitation de la cellule bipolaire) et centre OFF (le relâchement de glutamate au niveau centrale mène à une hyperpolarisation de la cellule bipolaire). Une cellule bipolaire à centre ON sera dépolarisé lorsque la lumière atteint le centre de son champ récepteur, et hyperpolarisé lorsqu'elle atteint la périphérie. Cet assemblage ON/OFF ainsi que les cellules horizontales permet un traitement préalable de l’information: il permet d’augmenter les contrastes de notre vision.
Les synapses des cellules bipolaires sont reliées aux cellules ganglionnaires de même type: une cellule bipolaire OFF sera relié à une cellule ganglionnaire OFF, et vice-versa. Cet conformation assure que l’information traitée par les cellules bipolaires est conservée. Il existe deux types majeurs de cellules ganglionnaires, les cellules P qui assurent une vision détaillée et la perception des couleur, et des cellules M qui ont pour objectif de discerner des formes et des mouvements grossiers. Leur distinction est faite grâce à leurs tailles: les cellules P ont d’étroits champ récepteurs, c’est-à-dire reliés à peu de cellules bipolaires, tandis que les cellules M en ont des importants. Contrairement aux photorécepteurs et aux cellules bipolaires, les neurones ganglionnaires ne se dépolarisent ou ne s’hyperpolarisent pas suite au relâchement de glutamate par la cellule bipolaire. Les cellules ganglionnaires vont avoir une fréquence de décharge de potentiel d’action plus ou moins importante selon le taux de glutamate relâché par les cellules bipolaire. Ainsi, si un faisceau de lumière touche la périphérie d’un champ récepteur d’une cellule bipolaire OFF, cela va mener à une dépolarisation de la cellule, qui va relâcher plus de glutamate, et la cellule ganglionnaire reliée à cette cellule bipolaire, excitée, va avoir une fréquence de décharge de potentiel d’action plus importante. Les cellules ganglionnaires possèdent des axones, un long fil nerveux, qui servent à propager le message nerveux. Ce message nerveux obéit une loi du “tout ou rien”, c’est-à-dire que le potentiel d’action généré par la cellule doit atteindre un seuil afin que l’axone envoie un signal compréhensible par le cerveau. Les axones des cellules ganglionnaires vont se regrouper afin de composer le nerf optique de l’oeil, qui propage l’information au cerveau.
De l’oreille au cerveau
Le cerveau, au lieu de l'oreille, est réellement responsable de la perception des sons. L'oreille est conçu pour convertir une onde mécanique (le son) en un signal nerveux. Notre système auditif procède comme un parcours qui conduit les vibrations sonores vers plusieurs milieux successifs. Ces milieux sont aérien dans l’oreille externe, solide dans l’oreille moyenne et liquide dans l’oreille interne.
Dans un premier temps, dirigées dans le conduit auditif externe par le pavillon, les vibrations auditives font résonner la membrane tympanique. Le rôle du tympan est de localiser la provenance des sons, de les capter puis de les amplifier. Comprenant le tympan et les osselets, c’est à dire le marteau, l’enclume et l’étrier, l’oreille moyenne assure le transfert des vibrations acoustiques jusqu’à l’entrée de l’oreille interne : la fenêtre ovale. Les vibrations sonores passent alors dans la rampe vestibulaire de la cochlée et stimulent l’organe de Corti situé à l’intérieur de celle-ci. Ses récepteurs détectent alors la fréquence et l’intensité des sons.
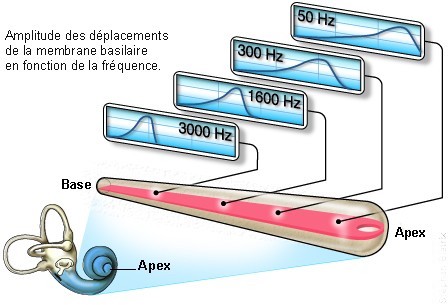
Schéma de fonctionnement du cochlée
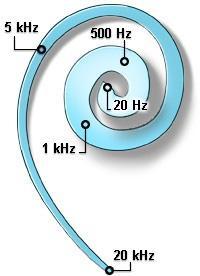
Schéma de fonctionnement du cochlée
Dans la cochlée, qui ressemble à une coquille d’escargot, les vibrations sonores font vibrer la membrane basilaire. Celle-ci va être déformé à différents endroits dépendant de la fréquence de l’onde sonore. La membrane basilaire contient des milliers de cellules ciliées. Ce sont ces cellules qui vont répondre à différentes fréquences. Les cellules sont placées comme dans un piano: les cellules qui sont activées par les fréquences les plus élevées sont localisées à la base du cochlée et plus on se rapproche de l’apex du cochlée plus les cellules réagissent à des fréquences basses.
Quand les cellules ciliées sont “poussées” assez loin suite à la déformation de la membrane basilaire, quand ces cellules sont stimulées, elles libèrent des ions afin de former un signal nerveux (ceci sera expliqué plus tard dans la partie sur le fonctionnement du système nerveux) pour être envoyé au cerveau où ce signal sera analysé.
Le transfert de l’information du cerveau aux muscles
Du cerveau au reste du corps
Comment un signal traverse le neurone: Le potentiel d’action
Le système nerveux est divisible en deux grandes parties:
- Le système nerveux central (SNC): constitué du cerveau et de la moelle épinière
- Le système nerveux périphérique (SNP): constitué de nerfs qui divergent du cerveau et de la moelle épinière; il permet au SNC de communiquer avec le reste du corps.
Lorsque un neurone est assez stimulé, il envoie un signal électrique le long de son axone pour le transmettre aux neurones voisins. Les neurones peuvent envoyer un seul signal uniforme en vitesse et tension. Par contre, un neurone peut varier la fréquence à laquelle il envoie ce message et c’est cela qui permet au cerveau d'interpréter différentes fréquences de manières différentes. C’est ce signal électrique qu’on appelle le potentiel d’action.
Le corps humain est électriquement neutre, mais cela n'empêche pas que certains endroits du corps soient plus ou moins chargé que d’autres. C’est cette différence de charge dans le corps que le corps lui-même exploite pour envoyer des messages nerveux. Le corps utilise des membranes comme barrières pour garder les charges positives et négatives séparées jusqu’au moment où le corps soit prêt à utiliser l'énergie que cette attraction crée. Si on compare un neurone à une pile, on prend la pile comme étant le neurone, la pile ne fait rien jusqu’à qu’elle soit connectée à quelque chose qui laisse les charges opposées de la pile bouger les unes vers les autres. Le neurone, lui, a besoin d’un évènement (l’activation des canaux ioniques mentionnés) pour rapprocher ces charges opposées les unes vers les autres.
Si on pense à un neurone du point de vue d’un électricien, un neurone a une tension et un courant:
- tension ou plutôt différence de potentiel électrique (en mV): mesure le potentiel d'énergie généré par la séparation des charges opposées; on appelle cette énergie le potentiel membranaire. Plus la différence entre la charge intracellulaire et la charge autour de la cellule est grande, plus la différence de potentiel électrique est grande et plus le potentiel membranaire est grand
- courant: c’est le flux d'électricité d’un point à un autre. Dans le cas d’une nerf, un courant est un mouvement d’ions positifs ou négatifs le long de la membrane qui offre une résistance à ce déplacement
Un neurone dans son état de repos est comme une pile dans son emballage qui n’est pas utilisée. Lorsqu’un neurone est dans son état de repos, l'intérieur de la cellule est chargé plus négativement comparé à l'extérieur de la cellule. La différence est appelée le potentiel de repos du neurone, mesuré à environ -70mV. En dehors de la cellule, il se trouve de nombreux ions sodium chargés positivement. La cellule, pendant son état de repos, est remplie d’ions potassium chargés eux aussi positivement; c’est donc les protéines dans la cellule qui sont chargées très négativement qui font la différence. En plus, il y a moins d’ions potassium à l'intérieur de la cellule qu’à l'extérieur. Donc un potentiel membranaire peut être créé; la cellule est polarisée.

Fonctionnement des pompes sodium-potassium
Des pompes sodium-potassium ont pour but de garder le potentiel membranaire à -70mV. Ces pompes sont des protéines localisées le long de l'axone d’un neurone, qui, pour chaque deux ions potassium qu’elle fait rentrer dans la cellule, trois ions sodium sortent.
Les pompes sodium-potassium ne sont pas les seules protéines le long de l'axone. Des grandes protéines appelées des canaux ioniques y sont aussi présentes. Il existe deux types principaux de canaux ioniques:
- canaux tensiodépendants qui s’ouvrent et se ferment quand un certain potentiel de membrane est atteint
- canaux chimio-dépendant qui s’ouvrent seulement quand un neurotransmetteur ou une hormone se fixe sur son récepteur
Lorsque ces canaux s’ouvrent, des ions se précipitent dans les canaux pour rétablir une neutralité électrique. Ce mouvement d'ions est la clé de tous les événements électriques dans les neurones: c’est ce qui nous permet d’agir, penser, sentir et éprouver des sentiments. Pour envoyer un message le long d’un axone, il faut une augmentation du potentiel membranaire assez élevée afin d’activer les canaux tensiodépendants qui sont les canaux les plus nombreux: cela crée un potentiel d’action.
Schéma du Fonctionnement des différents canaux
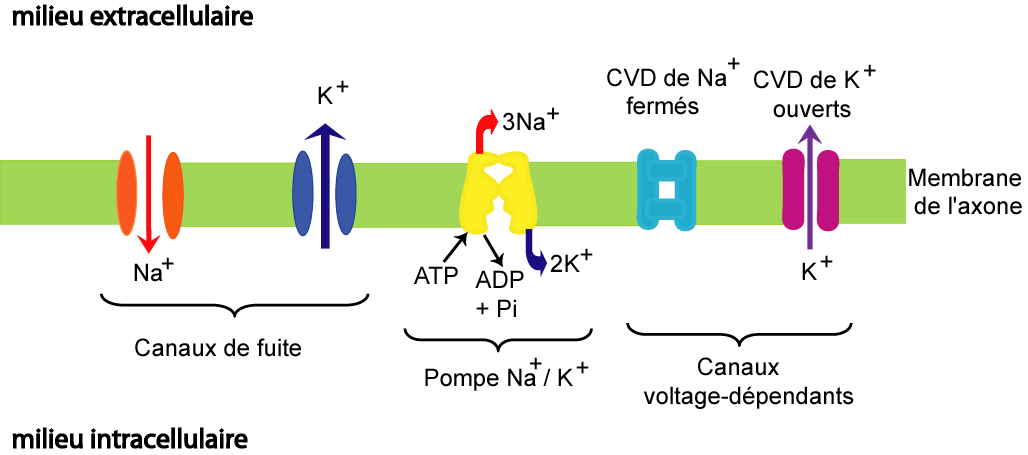
Voici comment se déroule le cycle d’un neurone afin qu’il puisse envoyer un message:
- Le neurone est en état de repos, tous les canaux ioniques sont fermés. Son potentiel membranaire est à -70mV.
- la concentration K+ est plus élevée à l'intérieur de la cellule qu’à l’extérieur
- la concentration de Na+ est supérieure à l'extérieur de la cellule qu’à l'intérieur
- Un stimulus est détecté.
- Des canaux sodiques sont ouverts.
- Des ions de sodium Na + rentrent donc dans la membrane et augmente la charge de celle-ci.
- Le stimulus doit être assez fort pour passer le seuil d’activation à un potentiel de membrane de -55mV.
- si le stimulus n’est pas assez fort pour passer ce seuil, le neurone retourne à son état de repos et attend un autre stimulus
- Par contre, si le seuil de -55mV est atteint, les canaux tensiodépendants sont activés.
- L’activation des canaux laisse un flux massif d’ions positifs rentrer dans la membrane du neurone. Ceci cause une dépolarisation massive: le potentiel de membrane monte à environ +40mV.
- Cette dépolarisation provoque une réaction en chaîne qui envoie le signal le long de l’axone. Étant donné qu’un neurone possède beaucoup de canaux tensiodépendants, quand quelques uns sont activés dans une région, le courant d’ions est assez fort pour changer la tension autour d’eux et donc active d’autres canaux tensiodépendants et ainsi de suite.
- Une fois que le signal atteint la terminaison axonale, le processus de repolarisation commence.
- Cette fois-ci, des canaux de potassium tensiodépendants s’ouvrent.
- Ils laissent des ions potassium K + sortir de la cellule afin de tenter de rééquilibrer les charges.
- Ceci provoque une hyperpolarisation: le potentiel de membrane passe brièvement à -75mV.
- Les canaux se ferment et les pompes potassium-sodium prennent le relais pour ramener la cellule à son potentiel de repos à -70mV.
Lorsqu'une partie d'un axone est en train d’envoyer un signal et que des canaux ioniques sont ouverts, il ne peut pas répondre à d’autres stimuli peu importe leur force. On appelle cela la période réfractaire qui existe afin de bloquer un signal essayant de passer dans l'axone dans deux directions à la fois. Elle dure environ 4 millisecondes.
Les neurones communiquent en un seul signal monotone: -70mV → +40mV → -75mV → -70mV → ...
Peu importe le stimulus, le signal reste le même et n’est pas changé. Par contre la fréquence change : Un stimulus faible a tendance à déclencher des potentiels d'action moins fréquents. Au contraire, des stimuli forts ont tendance à déclencher des potentiels d'action possédant des fréquences très élevées.
Les signaux varient également par leur vitesse: ils sont plus rapides dans les voies de nerfs qui réagissent à des choses telles que les réflexes, mais plus lents dans des endroits comme les vaisseaux sanguins.
Le potentiel d’action traverse les neurones généralement à une vitesse de 0.1 à 100 m.s-1. Le signal peut donc arriver n’importe où dans le corps en seulement quelques millisecondes.
Trois choses font varier la vitesse à laquelle un signal traverse un neurone:
- la température: des températures plus élevées augmentent le taux de diffusion des ions. Donc, plus la température est élevée, plus le signal traverse les neurones rapidement
- son diamètre: plus l’axone est large, moins il y a de résistance pour que le courant électrique traverse le neurone plus rapidement
- gaine de myéline: dans le système nerveux périphérique, ces gaines sont formées par les cellules de Schwann qui s’entourent le long de l'axone: ces cellules ne se touchent pas; il y a donc de petites lacunes entre la myéline appelées les nœuds de Ranvier.
Schéma d'un neurone myélinisé et son fonctionnement
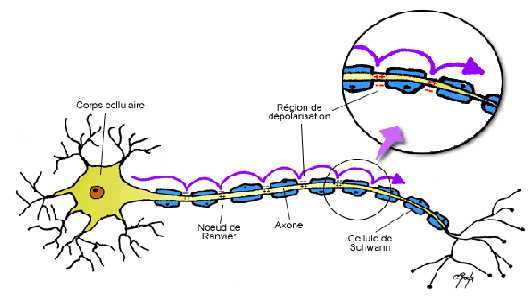 Ces gaines servent d’isolant électrique (elles empêchent la diffusion des ions). Le potentiel d'action peut donc sauter de grandes distances d’un nœud à un autre (1 mm) dans lesquelles les canaux tensiodépendants seront localisés (où la dépolarisation aura lieux), par un processus qui est appelé propagation saltatoire (du mot latin désignant “bond”). Cela augmente la vitesse de propagation de façon spectaculaire. Dans les neurones non-myélinsés, le potentiel d’action peut se déplacer à un vitesse maximale d’environ 1 m.s-1, alors que dans les neurones myélinisés, il peut se déplacer à plus de 100 m.s-1 Cependant, dans ce système nerveux central, les gaines de myléline sont formées par les oligodendrocytes. Une seule cellule est responsable de former la gaine de myéline mais son fonctionnement reste le même.
Ces gaines servent d’isolant électrique (elles empêchent la diffusion des ions). Le potentiel d'action peut donc sauter de grandes distances d’un nœud à un autre (1 mm) dans lesquelles les canaux tensiodépendants seront localisés (où la dépolarisation aura lieux), par un processus qui est appelé propagation saltatoire (du mot latin désignant “bond”). Cela augmente la vitesse de propagation de façon spectaculaire. Dans les neurones non-myélinsés, le potentiel d’action peut se déplacer à un vitesse maximale d’environ 1 m.s-1, alors que dans les neurones myélinisés, il peut se déplacer à plus de 100 m.s-1 Cependant, dans ce système nerveux central, les gaines de myléline sont formées par les oligodendrocytes. Une seule cellule est responsable de former la gaine de myéline mais son fonctionnement reste le même.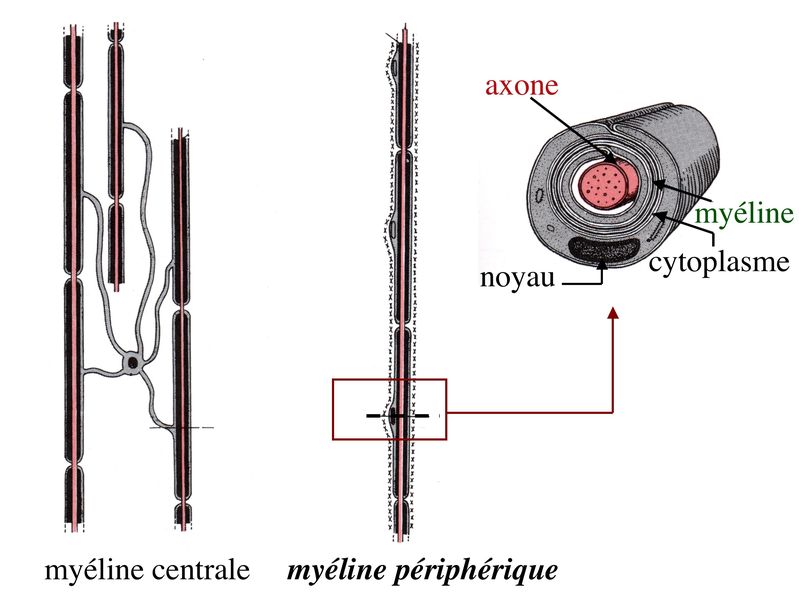
Schéma de la différence entre la myéline centrale et périphérique
Synapses
Cent fois plus petit que l'épaisseur d’une feuille de papier, plus nombreux en un seul individu que des grains de sable sur une plage: ce sont les synapses. La force et le but de l’existence des neurones résident dans leurs connexions. L'existence d’un neurone devient inutile s’il n'a personne à écouter, ni parler. Le mot synapse vient du mot grec pour dire “rejoindre”.
Lorsqu'un neurone envoie un message électrique vers l'extrémité d'un axone, ce signal atteint une synapse qui peut convertir le signal en un autre type de signal, puis passer ce même signal à un autre neurone. Ces connexions sont des exploits extraordinaires de la bio-ingénierie électrique. Ces connexions sont extrêmement nombreuses: le cerveau compte environ 100 milliards de neurones et chacun de ces neurones ont entre 1,000 et 10,000 synapses; soit environ un total de 100 à 1000 milliards de synapses uniquement dans le cerveau. Chaque synapse est comme un petit ordinateur capable d'exécuter plusieurs programmes simultanément et de changer ainsi que de s'adapter en réponse à des stimuli. Ils sont capables de renforcer ou d’affaiblir leurs connections en fonction de la fréquence à laquelle ils sont utilisés. Ce sont ces connections qui nous permettent d'apprendre et de mémoriser des choses. Tout est communiqué à travers ces signaux allant de l’euphorie, à la faim, à l’ennui et même au désir.

Les cellules nerveuses ont deux principales manières de communiquer les unes avec les autres en fonction de la vitesse à laquelle l’information a besoin de se déplacer. L’information peut se déplacer de deux façons:
- électriquement: un courant d’ions passe directement du cytoplasme d’une cellule à l’autre à travers de petites “fenêtres” appelées jonctions communicantes. Le signal n’est jamais converti. L’avantage de ce signal est qu’il est très rapide, mais il n’est pas précis: une seule synapse qui envoie un signal électrique peut activer des milliers d’autres cellules. C’est pratique dans le cœur par exemple, où il est nécessaire que toutes les cellules cardiaques soient stimulées en même temps. Si tous les neurones utilisaient ce type de signal pour communiquer, ils seraient toujours activés, ce qui ferait que tous les organes et tissus soient stimulés constamment, provoquant la mort par épuisement.
- chimiquement: utilisé plus souvent comme méthode de communication entre les neurones, cette méthode est plus lente, mais elle est plus précise. Ce type de communication utilise des neurotransmetteurs qui voyagent à travers la fente synaptique pour délivrer le message. Le message est donc converti d’électrique à chimique puis à l’électrique lorsque ce signal chimique atteint le neurone récepteur.
Au niveau des synapses, les signaux peuvent être modifiés, amplifiés, ou inhibées soit immédiatement ou sur une période de temps:
- potentialisation: renforcement d'une synapse
- accoutumance: la synapse diminue sa réponse à un stimulus commun
- sensibilisation: une réaction à un stimulus entraine d'autres synapses à être plus sensibles aux réactions
Le neurone présynaptique est le neurone qui envoie le signal. Ce neurone transmet un message à travers le terminal présynaptique généralement aux bornes des axones qui détiennent pleins de petites vésicules synaptiques chacun avec des milliers de neurotransmetteurs.
Le neurone post-synaptique est le neurone qui reçoit le signal du neurone présynaptique. Cette cellule reçoit les neurotransmetteurs dans une région du neurone où des récepteurs sont présents: habituellement sur les dendrites ou sur le soma de la cellule. Ces deux neurones ne se touchent jamais pour communiquer; il y a un petit écart entre les deux neurones appelés une fente synaptique séparant les deux de 50 nm.
Dans les synapses chimiques, les signaux ne sont pas transmis directement entre les neurones comme dans les synapses électriques.
La communication dans une synapse chimique implique la transmission, diffusion et réception de neurotransmetteurs. Cela résulte en une communication unidirectionnelle entre les neurones.
Quand un potentiel d’action atteint le bout d’un axone, c’est-à-dire la synapse, des canaux de calcium tensiodépendants s'activent pour ouvrir et libérer le calcium dans le cytoplasme du neurone. Ce flux de charges d’ions de calcium provoque le fusionnement des vésicules synaptiques avec la membrane cellulaire pour libérer les neurotransmetteurs contenus dans les vésicules dans la fente synaptique. Ces neurotransmetteurs vont ensuite se lier à des récepteurs sur le neurone post-synaptique. Le neurone présynaptique a donc converti le signal électrique en signal chimique.
C’est maintenant au tour du neurone post-synatique de reconvertir le signal chimique en potentiel d’action c’est-à-dire en signal électrique. Lorsque les neurotransmetteurs relâchés par le neurone présynaptique se lient aux récepteurs, cela provoque l’ouverture de canaux ioniques. En fonction du neurotransmetteur qui s’est lié à ces récepteurs, le neurone peut être excité ou inhibé.
- les neurotransmetteurs excitateurs dépolarisent le neurone post-synaptique en provoquant l'intérieur de celui-ci à devenir chargé plus positivement. Ce changement le rapproche du seuil de son potentiel d’action.
- les neurotransmetteurs inhibés hyperpolarisent le neurone post-synaptique en le rendant chargé encore plus négativement, l’éloignant de son seuil d’activation. Non seulement ce type de neurotransmetteur empêche des messages d'être envoyé, mais en plus rend le neurone plus difficile à exciter
Un neurone peut avoir des milliers de synapses. Ainsi, la probabilité qu’un neurone développe un potentiel d’action dépend de la somme de toutes les excitations et inhibitions dans la région autour des récepteurs de ce neurone. Notre corps produit une centaine de différents types de neurotransmetteurs qui occupent des fonctions différentes. Les neurotransmetteurs ne restent pas collés à leurs récepteurs pendant plus de quelques millisecondes. Ils sont ensuite soit recyclés (réabsorbés par le neurone présynaptique; détruits par des enzymes dans la fente synaptique, sortent de la synapse par diffusion) soit ils se dégradent.
 Exocytose
Exocytose
Le déroulement de l’action
La production du mouvement: la contraction musculaire
La production d’un mouvement, soit une action, est une des étapes du temps de réaction. Le temps de réaction dépend en partie du muscle sollicité pour réaliser l’action. On parle de contraction musculaire lorsque le muscle en question reçoit le potentiel d’action, évoqué précédemment, qui déclenche un stimulus et donc la production de nombreuses réactions chimiques aboutissant à cette contraction musculaire.
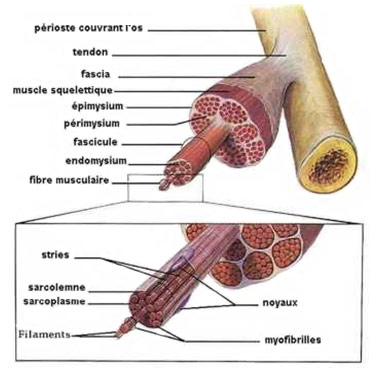
Anatomie d'un Muscle
Il faut savoir qu’un muscle est un assemblage de plusieurs parties. Le schéma suivant montre la composition d’un muscle:
Un muscle est composé de multiples faisceaux musculaires comportant chacun plusieurs fibres musculaires. Afin de créer un mouvement, un stimulus est transféré vers une de ces fibres musculaires dans le muscle concerné. Cette transmission se situe dans une portion de la membrane cellulaire appelée plaque motrice comme montrée ci-dessous.
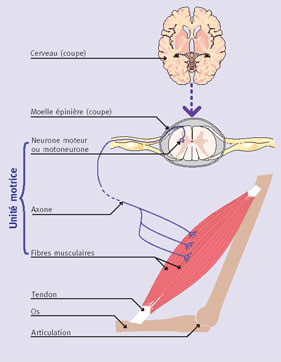
Connection du Muscle au reste du corps
Chaque fibre musculaire est en fait constituée de plusieurs petits filaments eux-mêmes comportant des cellules musculaires encore plus petits appelées myofibrilles. Suivant le transfert du stimulus vers la fibre musculaire, il y a une libération de molécules d’acétylcholine dans la fente synaptique. Ces molécules se fixent ensuite sur le sarcolemme qui s’agit d’une enveloppe autour du fibre musculaire. Lorsque ces molécules se fixent dessus, le sarcolemme devient plus ductile et va donc permettre la transmission d’ions de sodium et de potassium vers les myofibrilles. Ces ions sont nécessaires pour la contraction des muscles, car ils assurent la transmission des signaux nerveux au niveau des membranes cellulaires.
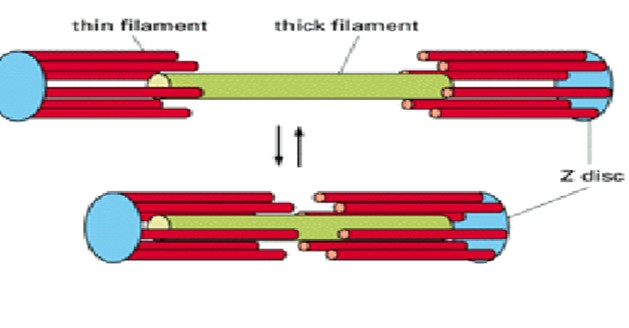
Connection du Muscle au reste du corps
Le stimulus généré va provoquer une contraction des filaments d’actine. Cette contraction est rendue possible grâce à une famille de protéines motrices spécifiques appelées les myosines. Ces filaments vont glisser entre les filaments de myosine avec l’aide de l’adénosine triphosphate (ATP), qui s’agit d’une cellule utilisée pour conserver toute l’énergie nécessaire pour le travail musculaire. L’ATP se situe également dans la fibre musculaire.
La fibre musculaire continuera à être excitée tant qu’il se trouve de l’acétylcholine dans la fente synaptique. L’acétylcholine est un neurotransmetteur permettant le passage de l’influx nerveux à travers la fente synaptique. Elle ne peut être détruite seulement par l’acétilcholinérase (enzyme catalysant).
Notre muscle est mis à l’effort lors d’une activité physique; par exemple, la jambe qui se contracte pour appuyer sur le frein, ou la main pour changer de vitesse dans une voiture. Tout ce processus provoque la contraction et ensuite la décontraction d’un muscle et déclenche l’alimentation de la fibre musculaire grâce aux nutriments stockés.